Bearcam is back for 2021, and while it’s still very early in the season several bears—including Grazer, Holly, and their yearlings—have made an appearance. As Rangers Naomi Boak and Lian Law discussed with me during our Welcome to Bearcam live chat, there are many fascinating storylines to follow this year. At the risk of offering a shameless plug, my book, The Bears of Brooks Falls, explores many of those stories too.
A dedicated book club has sprung up to discuss the book. At the end of each meeting, participants answer one question: If you could ask the author anything, what would be? Below, I’m happy to answer those questions. If you are interested in joining the book club for their next discussion on June 19 via Zoom, please sign up.
Questions from the club’s discussion of Part One: Creation and Discovery (May 29, 2021)
Can you clarify WHY there used to be fewer bears at the falls? In the past, were they hazed away? Did they stay away from the falls because anglers were given priority there?
In Part One of my book, I discuss the events that led to the proclamation of Katmai National Monument in 1918 and the monument’s evolution into one of the largest national parks in the United States. Bears were not a major tourist attraction at Brooks River until long after Brooks Lodge was established. It wasn’t because anglers were given priority. It was because the bear population was much smaller than today. The national monument was expanded in 1931 to include areas such as Brooks River to protect habitat for wildlife like bears, but:
By all accounts, few bears used the river when Brooks Lodge first opened for business in 1950. Bears and any type of bear-management activities were absent from the reports of the first rangers stationed at Brooks Camp. Ranger Russell Todd, for example, never saw a bear on foot in the summer of 1954. The presence of people alone was apparently enough of a deterrent to displace bears from the river except at night. In 1957, biologists conducting salmon research at Brooks River for the US Fish and Wildlife Service reported bears “loudly evident” every night during September at the salmon- counting weir strung across the head of the river.
How many bears lived within the monument at that time remains an open question, but it was likely not many. The population may even have been at a nadir, the result of decades of heavy hunting pressure near the monument and, I suspect, the lingering effects of the 1912 eruption. After a two- summer biological investigation of the monument in 1953 and 1954, Victor Cahalane reported: “It is impossible to make even a rough estimate of the population of bears in Katmai National Monument.” Yet he tried. According to his and other anecdotal sightings, including one from a pilot who claimed to have seen 60 bears along Savonoski River in early September 1954, Cahalane ventured that about 200 bears lived in the monument.
Steady levels of salmon and a reduction in hunting pressure outside the monument were probably the main factors that allowed the area’s bear population to slowly increase, but at Brooks Camp people inadvertently helped accelerate the bears’ use of the river. By the end of the 1960s, a small and growing contingent of bears had become accustomed to the easy access to unsecured food at nearby garbage dumps, the lodge’s burn barrels, and unsecured supplies. By the mid- 1970s, Brooks Camp had become well known as a place to find at least a few bears, and several had begun to fish in the river during the day when people were active. (Pg. 172-173, The Bears of Brooks Falls)
I will add that over the last 40 years, salmon runs in the Naknek River watershed have been quite strong and that, perhaps more than anything else, has allowed the bear population to increase in the park. Additionally, during much of that time, park staff management have emphasized minimizing bear-human conflicts. The experience of cubs that accompanied their mothers to Brooks River may now consist largely of relatively benign contacts with people. This probably allowed the number and proportion of adult bears tolerant of people to increase.
It sounds, from Mike’s description [in Chapter 3 — Ramble], that the outlet of Brooks Lake into Brooks River is pretty shallow. Could global warming threaten the snowfall on the mountains, dropping the level of the lake and halting the flow of the river? If so, could that be a risk in the near future?
Although I can only provide a speculative answer, and while Lake Brooks will be affected by a warmer atmosphere, its water flow may not change appreciably. Lake Brooks occupies a deep basin that is almost completely below the water table of the surrounding land. There are no glaciers in its headwaters, unlike nearby Naknek Lake, so it’s already adapted in a sense to a hydrology that is highly influenced by annual precipitation. Snowmelt is only one influence. After most of the snow melts from the watershed in late spring, then summertime rain seems to have the biggest influence on water levels in the lake. Wetter summers can raise lake levels more than a foot compared to dry summers. Importantly, much of its water is sourced from spring-fed streams and springs under the lake surface. So, even during drought years, the lake basin experiences some recharge.
Climate change is certainly altering Katmai’s landscape, both the land and water. In 2019, we saw the impacts of a very hot, dry summer on Brooks River. Water levels were quite low and water temperatures were quite hot during an early July heat wave that year. However, water continued to flow through the river, albeit at a reduced level.. That’s just one year, though. By the end of the century—especially if we don’t get our act together and reduce our greenhouse gas emissions as quickly as possible—the summer of 2019 will be one of the coolest of this century. Droughts and heat waves may become the norm in Katmai. For more information on the present and future of Katmai’s climate, please see chapter 17 of The Bears of Brooks Falls.
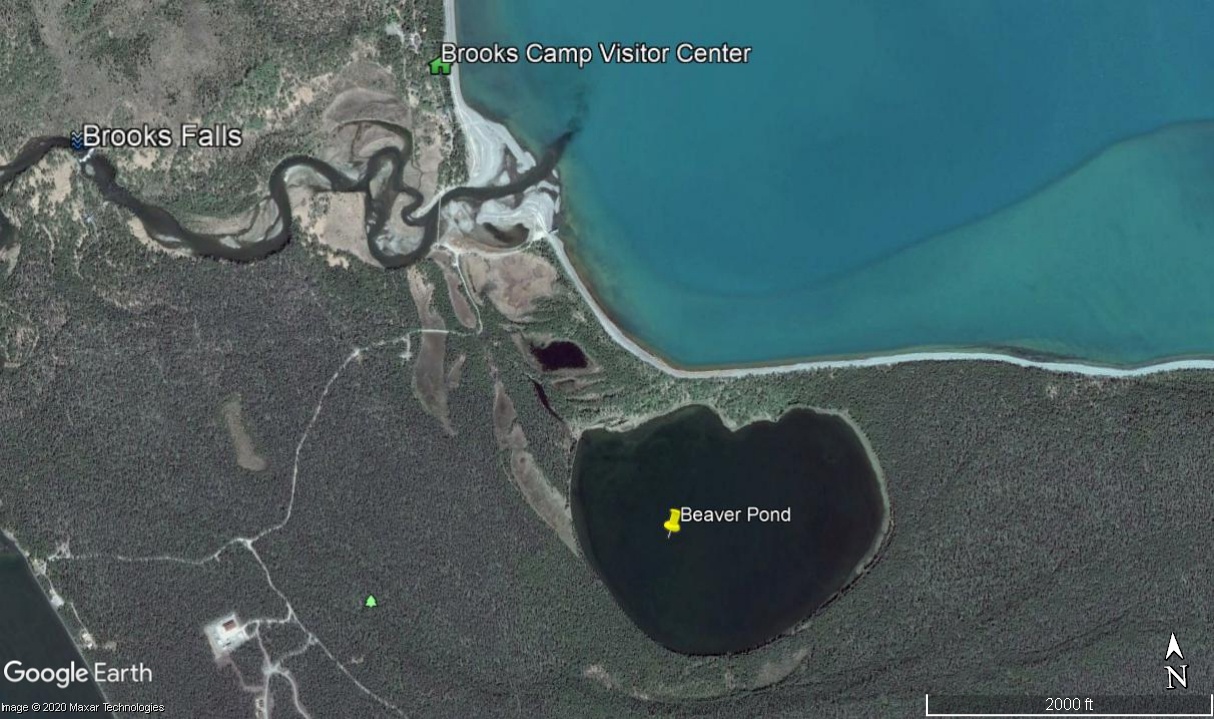
Can you clarify the distance from Brooks River to Margot Creek? Is it common — or uncommon — to see “our” cam bears at Margot Creek?
The shoreline of Naknek Lake between Brooks River and Margot Creek is about 13 miles, well within a day’s walk for a brown bear. If a bear takes a few shortcuts through the forest, then the walk is closer to 10-12 miles.

Several identifiable Brooks River bears use Margot Creek in August including 402, 435 Holly, 480 Otis, and 856. I would not be surprised if there are others well. But, salmon are dispersed widely in central Katmai in August when bears fish at Margot Creek. Unlike early summer and early fall when Brooks River is the only place to fish, bears have many other alternatives to Margot Creek in mid summer so not all Brooks River bears need to go there.
Can you talk about your research process? The book draws on your personal experience — but it clearly draws on extensive research, too.
When I began drafting the manuscript, I thought most of my research was finished since I had to study a lot to prepare programs and talk about bears when I was a park ranger at Katmai and through my current job at explore.org. That head start was helpful but not thorough enough. It was merely the foundation to build upon.
As I wrote, I wanted to be sure that my facts and conclusions were backed up by personal observations, experience, and the best available science. While working on the manuscript, I probably spent half my time reading research and half my time writing.
I began writing each chapter by outlining it. Then after I established what I wanted to write about and the stories that would add depth to the facts, I read or skimmed through the relevant books and scientific papers that I saved previously to establish the basic facts that I wanted to include and confirm what I thought I knew. This led me down many rabbit holes. I probably read dozens of papers for some chapters, especially Chapter 4 on hibernation. Tracking down specific facts and, hopefully, ensuring that I represented them accurately in the book was a tedious yet necessary task. Readers deserve no less.
Not a bear question, but a question for you as an author: What did it hurt to leave out of the book? What did you have to omit that you wish you’d been able to keep?
Quite a lot, actually. For example, I drafted chapters on glaciation and the evolution of Brooks River, but after consulting with an editor I decided to cut those. They weren’t a great fit for the narrative I tried to build. I also wanted to include the story of Holly adopting a yearling 503 in 2014 but couldn’t find the right place for it when I outlined the book. I considered using that story as the framework for Chapter 5: Family, but since adoption in bears is so uncommon I thought it best to focus on a bear whose maternal experiences were engaging yet more typical. That’s how I settled on 273 and her cub for Chapter 5. I’m happy with the final result of that chapter, yet I still wish I had found a way for Holly to be a part of it.
Questions from the club’s discussion of Chapter 6: Mating Season (June 6, 2021)
What if a female [bear] doesn’t want to mate? How much “say” does she have in the decision?
The female bear can’t control estrus or the signals that indicate to males that she is in estrus. However, female bears seem to have a lot of say in the timing of copulation. Although male bears are much larger than females, I’ve never seen a male bear force himself on a female bear. Instead, he doggedly follows her until she is ready to accept his advances. I also wonder if prolonged courtship can provide female bears with the chance to shed a suitor that they do not prefer. As I write in the book, a bear’s sense of smell is so powerful that a female can’t hide from a male. But, since mating opportunities are so limited for males, it’s not uncommon for more than one male to catch the scent of an estrous female. A prolonged estrus cycle coupled with a lengthy courtship could increase competition between males—an unconscious way for her to attract the most “fit” mate.
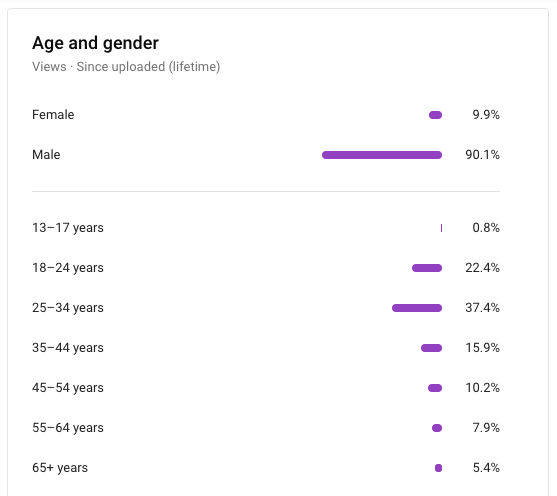
What is the ratio of males/females at Brooks River?
It hovers near 50:50, but last year there were more females than males. Because large adult male bears occupy the most productive fishing spots at Brooks Falls, it can sometimes seem like there are more males on the river than females. In July 2020 park bear monitoring staff identified slightly more female bears than males (29 adult females, 22 adult males, 14 subadult females, 11 subadult males).
Can you talk a bit about inbreeding? It seems like a lot of the bears we see mating are likely related to each other…
There’s only one confirmed case (through DNA analysis) of consanguineous couplings (inbreeding) between related bears at Brooks River.
24 BB was a very dominant male bear at Brooks River from the late 1990s through 2007. He was the equivalent of 856 during that time, and because of his dominance few bears would ever challenge him for fishing spots or for access to estrus females. BB sired a litter with the female 209. Bear 402, who still uses Brooks River, was one of the cubs from that litter born in 1998. 24 BB then sired a litter with 402. The offspring from the 402/24 relationship were weaned by 402 and identified as independent bears, but have not been seen in many years. I should note that this is common among subadult bears and their absence may not be reflective of interbreeding between a father bear and a daughter bear.
The limited DNA analysis of bears in 2005-2007 did not document any litters from a mother/son relationship. I think it’s unlikely that a bear could mate with its mother for a couple of reasons. 1. Male bears compete for the opportunity to mate with females and a larger, more dominant male would certainly outcompete a younger male bear for access. So while a young male bear is mature enough to mate around age 6, he’s still quite small compared to older males. 2. Young male bears often disperse away from their mother’s home range, and consequently their ranges as adults might not overlap. Mother bears remember who their offspring are too, and mom is often intolerant of the approach of her former cubs (we sometimes see a mother charge her former cubs, even years after family breakup, almost as if she is saying, “I told you to leave. Now stay away”).
Katmai’s brown bear population is quite large and robust. About 2,200 bears were estimated to live wholly or partly within Katmai National Park and Preserve in 2007. Although, we don’t know its true frequency, inbreeding between bears is probably uncommon here since the population is so large.
Why do mating males want to keep females in sight? It seems like all this following females around would distract males from eating and getting fat.
Courtship between bears isn’t always a prolonged process. In fact, sometimes bears couple soon after meeting. Potential male suitors, therefore, need to guard access to their prospective mates, lest they lose a rare mating opportunity.
The pursuit of mating opportunities certainly distracts male bears from other life tasks like fishing for salmon. I remember one July when 856 seemed like he didn’t stop courting females for the entire month. While the other males at the river got their fill of fish, 856 fished only occasionally because he was more interesting in reproduction. Near the end of July, he looked well muscled from the exercise of the pursuit but looked as though he had little body fat.

856 often spends a lot of time courting females in early summer and less time fishing compared to many other adult bears. He can afford to do so because his high level of dominance provides access to fishing spots wherever he goes.
“Survival of the fittest” is often thought to refer to athletic fitness or survival instincts, when it is more accurately framed in terms of reproductive fitness. Perhaps the male bears who have the energy reserves and stamina to court female bears for long periods of time with little food are the most reproductively fit. It’s also important to consider that the bears’ mating season ends in early summer, just when food becomes plentiful in Katmai, so a male who doesn’t eat much in June has ample opportunities to make up for it during the next few months.
Questions from the book club’s discussion of Chapter 14: Boundaries (June 12, 2021)
Is there any research showing how reduced attendance during the 2020 pandemic affected the salmon and/or the bears?
As far as I know, there’s nothing publicly available yet. However, biologists at Katmai National Park expanded the bear-monitoring program last year to collect data that might help answer that question. It was an unexpected research opportunity to observe bears at Brooks River at a time of year when typically it is loaded with people.
Certainly the lack of people at the river in 2020, especially when the camp remained closed to the public, allowed bears more space to fish. The greatest influence on the distribution of bears last year, though, was salmon. The record run of sockeye salmon was overwhelming and it provided bears with ample feeding opportunities throughout the river. In a year with fewer fish, I don’t think we wouldn’t have seen bears using the lower river in early summer as much as they did in 2020, no matter how few people visited.
The bears at Brooks are perhaps more human-habituated than other bears. And yet, as 854 Divot’s story proves, they do wander outside the boundaries of the park, where they will encounter humans who don’t operate according to park rules. Can you offer some reassurance — or some insight — about how their human habituation might affect their fate outside park boundaries?
Habituation at Brooks River provides a bear with advantages. It allows access to parts of the river that may otherwise be off limits if the bear isn’t tolerant of people. At Brooks River, people are especially tolerant of bears too through both attitude and regulations designed to protect bears.
Outside the park, they may not encounter the same tolerance. Having a bear prowling outside your cabin at Brooks Camp is one thing. Having it do so near your children and pets is another.
If a habituated bear wanders into King Salmon, for example, its tolerance for humans may lead it to temptation in the form of unsecured food and trash. A habituated bear could more easily become conditioned to seek human foods in that situation. Bears encounter much greater risks around people in those places than they do at Brook Camp.
Some biologists I’ve spoken to speculate that habituation could be context specific. That is, a bear might be able to learn that people in one location are tolerant while people in another location are dangerous. I think this is plausible but I’m not yet convinced it works that way for most bears. Further research is needed.











![Screen shot of youtube comment. Text says, "Gladhe 1 month ago He protecc He attacc He most importantly Eat big mac!]](https://fitznaturalist.com/wp-content/uploads/2021/01/screen-shot-2021-01-22-at-9.03.55-pm-1-e1611675551409.png)













_3_05022018")
_4_05022018")











 Curious to know more about its travels here, I followed the tracks along the edge of the river. The cougar followed the same general path I would have to move upstream; it stuck to the mud and driftwood on the edge of the willows. From the additional tracks I was able to find, the cougar continued along the riverbank for another hundred yards before I lost the trail in the adjacent thicket.
Curious to know more about its travels here, I followed the tracks along the edge of the river. The cougar followed the same general path I would have to move upstream; it stuck to the mud and driftwood on the edge of the willows. From the additional tracks I was able to find, the cougar continued along the riverbank for another hundred yards before I lost the trail in the adjacent thicket.









{kind=link}
{kind=link}